

Volume 7, Issue 2 (December 2021)
Elderly Health Journal 2021, 7(2): 79-83 |
Back to browse issues page



Download citation:
BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:



BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:
Soleimani H, Rezvani M E, Hafizi-Barjin Z, Esmaeilidehaj M, Zaremehrjerdi F. Effects of Chlorogenic Acid on Epileptic Behavior and mRNA Expressions of Brain Derived Neurotrophic Factor in the Brain of Aged Rats. Elderly Health Journal 2021; 7 (2) :79-83
URL: http://ehj.ssu.ac.ir/article-1-125-en.html
URL: http://ehj.ssu.ac.ir/article-1-125-en.html
Hossain Soleimani 
 , Mohammad Ebrahim Rezvani *
, Mohammad Ebrahim Rezvani *  , Zainab Hafizi-Barjin , Mansour Esmaeilidehaj , Fatemeh Zaremehrjerdi
, Zainab Hafizi-Barjin , Mansour Esmaeilidehaj , Fatemeh Zaremehrjerdi
, Mohammad Ebrahim Rezvani * , Zainab Hafizi-Barjin , Mansour Esmaeilidehaj , Fatemeh Zaremehrjerdi
Department of Physiology, School of Medicine, Shahid Sadoughi University of Medical Sciences, Yazd, Iran , erezvani@yahoo.com
Full-Text [PDF 1297 kb]
(449 Downloads)
| Abstract (HTML) (2221 Views)
.png)

Figure 1. The anti-seizure effects of different doses of CA in compared to control rats. GTCS: generalized tonic clonic seizure, *p < 0.05 and **p < 0.01when compared to PTZ + Veh

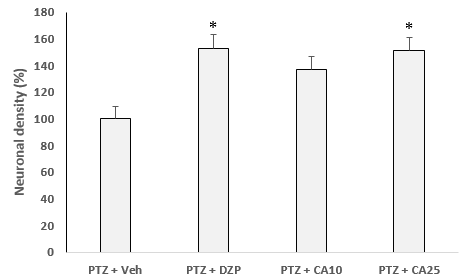
Figure 2. Histological changes and morphometrics of different doses of CA in compared to control rats. GTCS: generalized tonic clonic seizure, *p < 0.05 and **p < 0.01when compared to PTZ + Veh
Full-Text: (554 Views)
Effects of Chlorogenic Acid on Epileptic Behavior and mRNA Expressions of Brain Derived Neurotrophic Factor in the Brain of Aged Rats
Hossain Soleimani.1, Mohammad Ebrahim Rezvani *1, Zainab Hafizi-Barjin 1, Mansour Esmaeilidehaj 1, Fatemeh Zaremehrjerdi 1
Article history
Received 7 Oct 2018
Accepted 19 Oct 2021
A B S T R A C T
Hossain Soleimani.1, Mohammad Ebrahim Rezvani *1, Zainab Hafizi-Barjin 1, Mansour Esmaeilidehaj 1, Fatemeh Zaremehrjerdi 1
- Department of Physiology, School of Medicine, Shahid Sadoughi University of Medical Sciences, Yazd, Iran
Article history
Received 7 Oct 2018
Accepted 19 Oct 2021
A B S T R A C T
Introduction: The present study was conducted to evaluate the effect of chlorogenic acid (CA) and Diazepam (DZP) on epileptic complication that induced by repetitive intra-peritoneal injections of Pentylenetetrazle (PTZ) in aged rats.
Methods: Twenty-four month-old male Wistar rats (age > 12 months, 300-350 g) were divided in 4 experimental groups. Animal in control group (PTZ + Vehicle) received only PTZ. Animal in treated groups (PTZ + DZP, PTZ + CA10 and PTZ + 25) received diazepam 2 mg/kg, CA 10 mg/kg, or CA 25 mg/kg. All drugs injection were performed 30 min prior to each PTZ injection. Epilepsy was induced by injection of subconvulsive dose of PTZ every other day until the rats were completely kindled or epileptic. After each PTZ injection, animal was monitored for 40 min and epileptic behaviors were scored. At the end of the study, rats were sacrificed and the brains removed for evaluation of histological changes and Brain Derived Neurothrophic Factor (BDNF) gene expression.
Results: CA at dose of 25 mg/kg reduced percent of generalized tonic-clonic seizure (GTCS) in aged rats (24%) in compared to control group (50%) (p < 0.05). The latencies to the start of GTCS were decreased in both dose of CA (p < 0.05). Also, the percent of survived neurons in rats treated with CA (154%) were significantly higher relative to that of control animals (100%) (p < 0.05). The mRNA levels of BDNF significantly increased in CA treated rats (p < 0.05).
Conclusion: Hence, these findings revealed that CA have antiepileptic, neuroprotective and trophic effects in aged rats. CA can protect aged brain from deteriorative processes and save neurons during epilepsy in rats.
Keywords: Aging, Rat, Epilepsy, Chlorogenic Acid, Brain Derived Neurotropic Factor
Methods: Twenty-four month-old male Wistar rats (age > 12 months, 300-350 g) were divided in 4 experimental groups. Animal in control group (PTZ + Vehicle) received only PTZ. Animal in treated groups (PTZ + DZP, PTZ + CA10 and PTZ + 25) received diazepam 2 mg/kg, CA 10 mg/kg, or CA 25 mg/kg. All drugs injection were performed 30 min prior to each PTZ injection. Epilepsy was induced by injection of subconvulsive dose of PTZ every other day until the rats were completely kindled or epileptic. After each PTZ injection, animal was monitored for 40 min and epileptic behaviors were scored. At the end of the study, rats were sacrificed and the brains removed for evaluation of histological changes and Brain Derived Neurothrophic Factor (BDNF) gene expression.
Results: CA at dose of 25 mg/kg reduced percent of generalized tonic-clonic seizure (GTCS) in aged rats (24%) in compared to control group (50%) (p < 0.05). The latencies to the start of GTCS were decreased in both dose of CA (p < 0.05). Also, the percent of survived neurons in rats treated with CA (154%) were significantly higher relative to that of control animals (100%) (p < 0.05). The mRNA levels of BDNF significantly increased in CA treated rats (p < 0.05).
Conclusion: Hence, these findings revealed that CA have antiepileptic, neuroprotective and trophic effects in aged rats. CA can protect aged brain from deteriorative processes and save neurons during epilepsy in rats.
Keywords: Aging, Rat, Epilepsy, Chlorogenic Acid, Brain Derived Neurotropic Factor
Copyright © 2021 Elderly Health Journal. This is an open-access article distributed under the terms of the Creative Commons Attribution-Non Commercial 4.0 International License (http://creativecommons.org/licenses/by-nc/4.0/) which permits copy and redistribute the material just in noncommercial usages, provided the original work is properly cite.
Introduction
The incidence of epileptic disorders has a high frequency in the elderly. With increasing chronologic age, the significant changes in different brain structures occurring in epilepsy appear to progress in an age-related fashion compared to non-epileptic controls. These structural alternations reveal that elderly persons with epilepsy represent more sever cognitive deficiencies (1).
The population of older individuals (ages more than 65 years) will increase as time passed (1). Aging as a physiological process causes several persistence disturbances in the homeostatic mechanisms, accompanied by deficiency in learning and memory functions and cognitive decline. Two of the brain areas damaged by aging processes are the hippocampus and prefrontal cortex, which causes memory and learning disorders (2). During aging, these areas undergo some cellular and molecular changes that leads to learning, memory and cognitive dysfunctions.
There are many proteins as neurotrophic factors that regulate synaptic function and neuronal growth. Signaling of these neurotrophic factors are also severely affected in aging process and brain degenerative diseases which can be correlated with cognitive decline. Among these factors, Brain-derived neurotrophic factor (BDNF) and nerve growth factor (NGF) are the most important neurotrophic factors in preventing the degenerative and deteriorative disorders of brain that coincide with aging (3, 4).
Wide spectrum of antiepileptic drugs (AEDs) can lead to seizure suppression and are of great clinical importance because they deteriorate seizure severity and symptoms (5). However, seizure-induced neurodegeneration, drug-resistive epileptogenesis and their serious side effects are the main goals in treatment of epileptic diseases (6, 7).Therefore, investigation for complementary, alternative and novel drug from natural sources is essential to improve seizure and concurrently protect cognitive dysfunctions especially in elderly epileptic patients.
Generally, Chlorogenic acid (CA) as a polyphenolic compound, have the antioxidant, antidepressant, anti-inflammatory, analgesic and anti-apoptotic activities (8-10). However, the neuroprotective role of this natural occurred compounds has previously been reported (8). Recent studies indicated that phenolic acids such as chlorogenic acid exert anticholinesterase and cognition-improving effects and reduction of the pilocarpine-induced epilepsy (seizures) in mice (11, 12).
Based on these evidences, we conducted this study to determine the role of chlorogenic acid on brain structures and neurotrophic factors in aged epileptic rats.
Methods
Animals
Twenty-four month-old male Wistar rats were used in the present study. They were maintained at 25 ± 2 ºC with 12 h dark/light cycle and 40–70% humidity. The animals had free access to food and water. The rats were fed with a standard rat chow diet. Experiments were carried out after a week of acclimatization.
Chemicals
Pentylenetetrazole (PTZ) was purchased from Sigma-Aldrich (UK) and dissolved in sterile saline 0.9% on a weight/volume basis on the day of use. CA obtained from Tocris Bioscience (Bristol, UK) and dissolved in0.1% dimethyl sulfoxide (DMSO) in various concentrations that are described in the next section.
Induction and evaluation of PTZ kindling
The rats were randomly divided into 4 experimental groups with 6 animals in each. Rats in control group (PTZ + Veh) received 0.2 ml of 0.1% DMSO as vehicle 30 minutes before PTZ injection at the dose of 35mg/kg. Animal in treated groups (PTZ + DZP, PTZ + CA10 and PTZ + 25) received diazepam 2 mg/kg, CA 10 mg/kg, or CA 25 mg/kg respectively 30 minutes prior to 35 mg/kg PTZ (i.p.).
Subsequent to PTZ injections, animals were monitored for 40 minutes and epileptic behaviors were scored as follows: 0; no behavioral changes. 1; facial movements, ear and whisker twitching. 2; myoclonic convulsions without rearing. 3; myoclonic convulsions with unilateral forelimb clonus. 4; myoclonic convulsions with rearing. 5; loss of posture and generalized clonic-tonic seizures (13).
Histological survey
At the end of experiment, the brain of rats was removed fixed in neutral buffer formalin, processed and paraffin embedded as per the standard protocol. Tissue samples of all rats were sectioned coronally in 5µm thickness. Tissue staining with Hematoxyl-Eosin (H&E) was then performed. The live and normal neuronal cells were defined as round-shaped, cytoplasmic membrane-intact cells, without any nuclear condensation or distorted aspects. The normal neuronal cells in the hippocampal areas were seen at high magnification using light microscope (Zeiss, Germany) (14).
Quantitative Real Time - PCR
Tissue mRNA levels of BDNF were determined using quantitative real-Time polymerase chain reaction. RNA of each tissue sample of brain was extracted using RNA extraction kit (Cinna pure TMkit, Iran) according to the kit instruction. Efficiency and quality of RNA extraction were determined using 260/280 ratios of optical density from each sample (microplate reader, Epoch, England). Single strand cDNA was synthesized from 1µg of the extracted mRNA using the RevertAid™ First Strand cDNA Synthesis kit and random hexanucleotide primers (Thermofischer, USA). qRT-PCR was performed by using master mix containing SYBR green DNA dye (Amplicon, Denmark). The qPCR reaction contained 12.5 μl of the Master Mix, 10.5 μl of purified water, 1 μl of 10 μM primer set, and 1 μl of cDNA template. All values were normalized to RP13, a housekeeping gene with minimal variability. Primer sequences, annealing temperatures, and product sizes are presented in Table 1. Each gene was amplified for 40 cycles and ∆∆Ct method was applied to compare the relative gene expression.
Ethical considerations
All procedures were carried out according to the guidelines of Animal Ethics Committee of the Shahid Sadoughi University of Medical Sciences (Local ethical code: IR.SSU.MEDICINE.REC.1395.41).
Data analysis
GraphPad Prism software (version 8) was utilized for the statistical data analysis, and the obtained data were expressed in Mean ± standard error of the mean (SEM). One-way analysis of variance (ANOVA) and Tukey's posthoc test was used to find significant differences between all experimental groups. p <0.05 is considered statistically significant in all experiments.
Result
Anti-seizure effects
CA at doses of 25 mg/kg but not at dose of 10 mg/kg could significantly decrease the percent of generalized tonic-clonic seizures in PTZ-treated rats relative to control group (p < 0.05). Also, rats received both doses of 10 or 25 mg/kg exhibited lower GTCS latencies relative to control group (p < 0.05). Diazepam had a decreasing effect on the percent of GTCS and its latencies in PTZ treated rats when compared to those of control groups (p < 0.05). Anti-seizure effects of CA shown in figure 1.
Histological changes
H&E staining showed that repetitive convulsion cause to neuronal loss which is evident in Figure 2. A significant improvement in the neuronal densities in the CA1 region was observed in DZP or CA treated group in comparison to the control group. Also, as seen in figure 2, treatment of convulsive rats with DZP or CA at dose of 25 mg/kg lead to increase the density of neurons in the hippocampus of PTZ treated rats in compared to that of control rats.
Molecular findings
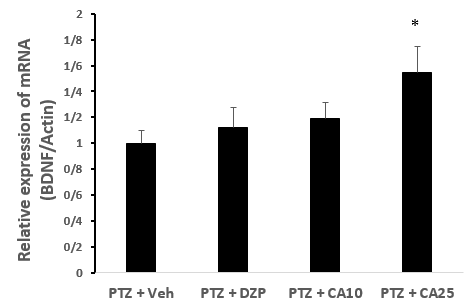
In order to evaluate the relative expression of BDNF in hippocampus of control and treated convulsive animals, the level of these neurotrophic factors was determined by semi-quantitative RT-PCR. Real-time PCR analysis exhibited that relative expressions of BDNF were increase in the hippocampus of DZP or CA at doses of 10 and 25 mg/kg injected rats (Fig. 3).
The population of older individuals (ages more than 65 years) will increase as time passed (1). Aging as a physiological process causes several persistence disturbances in the homeostatic mechanisms, accompanied by deficiency in learning and memory functions and cognitive decline. Two of the brain areas damaged by aging processes are the hippocampus and prefrontal cortex, which causes memory and learning disorders (2). During aging, these areas undergo some cellular and molecular changes that leads to learning, memory and cognitive dysfunctions.
There are many proteins as neurotrophic factors that regulate synaptic function and neuronal growth. Signaling of these neurotrophic factors are also severely affected in aging process and brain degenerative diseases which can be correlated with cognitive decline. Among these factors, Brain-derived neurotrophic factor (BDNF) and nerve growth factor (NGF) are the most important neurotrophic factors in preventing the degenerative and deteriorative disorders of brain that coincide with aging (3, 4).
Wide spectrum of antiepileptic drugs (AEDs) can lead to seizure suppression and are of great clinical importance because they deteriorate seizure severity and symptoms (5). However, seizure-induced neurodegeneration, drug-resistive epileptogenesis and their serious side effects are the main goals in treatment of epileptic diseases (6, 7).Therefore, investigation for complementary, alternative and novel drug from natural sources is essential to improve seizure and concurrently protect cognitive dysfunctions especially in elderly epileptic patients.
Generally, Chlorogenic acid (CA) as a polyphenolic compound, have the antioxidant, antidepressant, anti-inflammatory, analgesic and anti-apoptotic activities (8-10). However, the neuroprotective role of this natural occurred compounds has previously been reported (8). Recent studies indicated that phenolic acids such as chlorogenic acid exert anticholinesterase and cognition-improving effects and reduction of the pilocarpine-induced epilepsy (seizures) in mice (11, 12).
Based on these evidences, we conducted this study to determine the role of chlorogenic acid on brain structures and neurotrophic factors in aged epileptic rats.
Methods
Animals
Twenty-four month-old male Wistar rats were used in the present study. They were maintained at 25 ± 2 ºC with 12 h dark/light cycle and 40–70% humidity. The animals had free access to food and water. The rats were fed with a standard rat chow diet. Experiments were carried out after a week of acclimatization.
Chemicals
Pentylenetetrazole (PTZ) was purchased from Sigma-Aldrich (UK) and dissolved in sterile saline 0.9% on a weight/volume basis on the day of use. CA obtained from Tocris Bioscience (Bristol, UK) and dissolved in0.1% dimethyl sulfoxide (DMSO) in various concentrations that are described in the next section.
Induction and evaluation of PTZ kindling
The rats were randomly divided into 4 experimental groups with 6 animals in each. Rats in control group (PTZ + Veh) received 0.2 ml of 0.1% DMSO as vehicle 30 minutes before PTZ injection at the dose of 35mg/kg. Animal in treated groups (PTZ + DZP, PTZ + CA10 and PTZ + 25) received diazepam 2 mg/kg, CA 10 mg/kg, or CA 25 mg/kg respectively 30 minutes prior to 35 mg/kg PTZ (i.p.).
Subsequent to PTZ injections, animals were monitored for 40 minutes and epileptic behaviors were scored as follows: 0; no behavioral changes. 1; facial movements, ear and whisker twitching. 2; myoclonic convulsions without rearing. 3; myoclonic convulsions with unilateral forelimb clonus. 4; myoclonic convulsions with rearing. 5; loss of posture and generalized clonic-tonic seizures (13).
Histological survey
At the end of experiment, the brain of rats was removed fixed in neutral buffer formalin, processed and paraffin embedded as per the standard protocol. Tissue samples of all rats were sectioned coronally in 5µm thickness. Tissue staining with Hematoxyl-Eosin (H&E) was then performed. The live and normal neuronal cells were defined as round-shaped, cytoplasmic membrane-intact cells, without any nuclear condensation or distorted aspects. The normal neuronal cells in the hippocampal areas were seen at high magnification using light microscope (Zeiss, Germany) (14).
Quantitative Real Time - PCR
Tissue mRNA levels of BDNF were determined using quantitative real-Time polymerase chain reaction. RNA of each tissue sample of brain was extracted using RNA extraction kit (Cinna pure TMkit, Iran) according to the kit instruction. Efficiency and quality of RNA extraction were determined using 260/280 ratios of optical density from each sample (microplate reader, Epoch, England). Single strand cDNA was synthesized from 1µg of the extracted mRNA using the RevertAid™ First Strand cDNA Synthesis kit and random hexanucleotide primers (Thermofischer, USA). qRT-PCR was performed by using master mix containing SYBR green DNA dye (Amplicon, Denmark). The qPCR reaction contained 12.5 μl of the Master Mix, 10.5 μl of purified water, 1 μl of 10 μM primer set, and 1 μl of cDNA template. All values were normalized to RP13, a housekeeping gene with minimal variability. Primer sequences, annealing temperatures, and product sizes are presented in Table 1. Each gene was amplified for 40 cycles and ∆∆Ct method was applied to compare the relative gene expression.
Ethical considerations
All procedures were carried out according to the guidelines of Animal Ethics Committee of the Shahid Sadoughi University of Medical Sciences (Local ethical code: IR.SSU.MEDICINE.REC.1395.41).
Data analysis
GraphPad Prism software (version 8) was utilized for the statistical data analysis, and the obtained data were expressed in Mean ± standard error of the mean (SEM). One-way analysis of variance (ANOVA) and Tukey's posthoc test was used to find significant differences between all experimental groups. p <0.05 is considered statistically significant in all experiments.
Result
Anti-seizure effects
CA at doses of 25 mg/kg but not at dose of 10 mg/kg could significantly decrease the percent of generalized tonic-clonic seizures in PTZ-treated rats relative to control group (p < 0.05). Also, rats received both doses of 10 or 25 mg/kg exhibited lower GTCS latencies relative to control group (p < 0.05). Diazepam had a decreasing effect on the percent of GTCS and its latencies in PTZ treated rats when compared to those of control groups (p < 0.05). Anti-seizure effects of CA shown in figure 1.
Histological changes
H&E staining showed that repetitive convulsion cause to neuronal loss which is evident in Figure 2. A significant improvement in the neuronal densities in the CA1 region was observed in DZP or CA treated group in comparison to the control group. Also, as seen in figure 2, treatment of convulsive rats with DZP or CA at dose of 25 mg/kg lead to increase the density of neurons in the hippocampus of PTZ treated rats in compared to that of control rats.
Molecular findings
In order to evaluate the relative expression of BDNF in hippocampus of control and treated convulsive animals, the level of these neurotrophic factors was determined by semi-quantitative RT-PCR. Real-time PCR analysis exhibited that relative expressions of BDNF were increase in the hippocampus of DZP or CA at doses of 10 and 25 mg/kg injected rats (Fig. 3).
Figure 1. The anti-seizure effects of different doses of CA in compared to control rats. GTCS: generalized tonic clonic seizure, *p < 0.05 and **p < 0.01when compared to PTZ + Veh
Figure 2. Histological changes and morphometrics of different doses of CA in compared to control rats. GTCS: generalized tonic clonic seizure, *p < 0.05 and **p < 0.01when compared to PTZ + Veh
Figure 3. Relative expression of BDNF mRNA in different groups treated with CA in compared to control rats. GTCS: generalized tonic clonic seizure, *p < 0.05 when compared to PTZ + Veh
Discussion
Discussion
In the present study, we have examined whether the treatment with CA or DZP can attenuate neurodegeneration in hippocampus observed during seizures induced by PTZ in aged rats. Pretreatment of polyphenolic compounds can powerfully prevent the hippocampal neurodegeneration caused by destructive free radical species which is evident in the results (15). The distributions of the bioactive antioxidant compounds are not well understood, but then their bioavailability is a topic of considerable debate. CA is a common polyphenols found in a wide variety of food and beverages frequently consumed by humans such as olive oil, spices, wine, coffee, apples, cherries, plums, berries, and apricots (16). A broad literature searches on CA revealed that it exhibits many pharmacological effects including cardioprotective effects, lipid hyperoxidation inhibitory activity, anti-tumor and antioxidant activities (17, 18). In addition there are mounting evidences that regular dietary uptake of CA could fabricate a wide range of health benefits and reduce the risk of developing neurodegenerative diseases, which is evident from our results (19, 20). Oxidative stress during epilepsy induced by pilocarpine was also supported by studies showing beneficial effects of antioxidant compounds during seizure (21).
Epilepsy has been proposed to be associated with an imbalance in the excitatory (glutamate) and inhibitory amino acids (GABA) that play a crucial role in cognitive processes and neurodegeneration. Decreased GABA and neurotrophic factors such as BDNF level or increased glutamate can be lead to increased anxiety, aggression and reduced social reactive responses (22). In many of the experimental and human epileptic studies a decrease in the BDNF was reported (23, 24). The present study exhibited decreased level of BDNF in non-CA treated epileptic rats. Treatment with CA as well as standard drug diazepam significantly increased the level of BDNF in aged rats.
Conclusion
Our result in this study suggested that polyphenolic compound CA show antiepileptic and neuroprotective effects in aged rat epileptic model. CA can lead to attenuate epileptic behaviors and prevent the decreases in BDNF and NGF which occur during repetitive epilepsy. These findings disclose the protective role of CA as a dietary polyphenolics compound as useful neuroprotectiveagents in epileptic complications.
Study limitations
This study was performed in only aged Wistar rats and its findings cannot be extended to all species. Also, the lack of dose response study for obtaining the wide range of chlorogenic effects was a limitation of study.
Conflict of interests
The authors declare that they have no conflict of interest.
Acknowledgments
This article was derived from the M.sc thesis at Shahid Sadoughi University of Medical Sciences, Iran. We gratefully all people who assisted us in carrying out this study.
Authors’ contributions
Hossain Soleimani performed the laboratory work and writing the manuscript. Mohammad Ebrahim Rezvani designed and supervised the research work and the manuscript. Zainab Hafizi-Barjin contributed in laboratory technical, Mansour Esmaeilidehaj and Fatemeh Zaremehrjerdi advised this study and improve the statistical and histological analysis.
References
Epilepsy has been proposed to be associated with an imbalance in the excitatory (glutamate) and inhibitory amino acids (GABA) that play a crucial role in cognitive processes and neurodegeneration. Decreased GABA and neurotrophic factors such as BDNF level or increased glutamate can be lead to increased anxiety, aggression and reduced social reactive responses (22). In many of the experimental and human epileptic studies a decrease in the BDNF was reported (23, 24). The present study exhibited decreased level of BDNF in non-CA treated epileptic rats. Treatment with CA as well as standard drug diazepam significantly increased the level of BDNF in aged rats.
Conclusion
Our result in this study suggested that polyphenolic compound CA show antiepileptic and neuroprotective effects in aged rat epileptic model. CA can lead to attenuate epileptic behaviors and prevent the decreases in BDNF and NGF which occur during repetitive epilepsy. These findings disclose the protective role of CA as a dietary polyphenolics compound as useful neuroprotectiveagents in epileptic complications.
Study limitations
This study was performed in only aged Wistar rats and its findings cannot be extended to all species. Also, the lack of dose response study for obtaining the wide range of chlorogenic effects was a limitation of study.
Conflict of interests
The authors declare that they have no conflict of interest.
Acknowledgments
This article was derived from the M.sc thesis at Shahid Sadoughi University of Medical Sciences, Iran. We gratefully all people who assisted us in carrying out this study.
Authors’ contributions
Hossain Soleimani performed the laboratory work and writing the manuscript. Mohammad Ebrahim Rezvani designed and supervised the research work and the manuscript. Zainab Hafizi-Barjin contributed in laboratory technical, Mansour Esmaeilidehaj and Fatemeh Zaremehrjerdi advised this study and improve the statistical and histological analysis.
References
- Hernández-Ronquillo L, Adams S, Ballendine S, Téllez-Zenteno JF. Epilepsy in an elderly population: Classification, etiology and drug resistance. Epilepsy Research. 2018; 140: 90-4.
- Erdő F, Denes L, de Lange E. Age-associated physiological and pathological changes at the blood–brain barrier: a review. Journal of Cerebral Blood Flow & Metabolism. 2017; 37(1): 4-24.
- Mercado NM, Collier TJ, Sortwell CE, Steece-Collier K. BDNF in the aged brain: translational implications for Parkinson’s disease. Austin Neurology & Neurosciences. 2017; 2(2).
- Mariga A, Mitre M, Chao MV. Consequences of brain-derived neurotrophic factor withdrawal in CNS neurons and implications in disease. Neurobiology of Disease. 2017; 97: 73-9.
- Brunton L, Chabner B, Knollman B. Goodman and Gilman's the pharmacological basis of therapeutics. New York: McGraw-Hill; 1996.
- Bethmann K, Fritschy JM, Brandt C, Löscher W. Antiepileptic drug resistant rats differ from drug responsive rats in GABAA receptor subunit expression in a model of temporal lobe epilepsy. Neurobiology of Disease. 2008; 31(2): 169-87.
- Ali I, Chugh D, Ekdahl CT. Role of fractalkine–CX3CR1 pathway in seizure-induced microglial activation, neurodegeneration, and neuroblast production in the adult rat brain. Neurobiology of Disease. 2015; 74:194-203.
- Bravo L. Polyphenols: chemistry, dietary sources, metabolism, and nutritional significance. Nutrition Reviews. 1998; 56(11): 317-33.
- Basli A, Soulet S, Chaher N, Mérillon JM, Chibane M, Monti JP, et al. Wine polyphenols: potential agents in neuroprotection. Oxidative Medicine and Cellular Longevity. 2012; 2012: 1-14.
- Ozcan T, Akpinar-Bayizit A, Yilmaz-Ersan L, Delikanli B. Phenolics in human health. International Journal of Chemical Engineering and Applications. 2014; 5(5): 393-6.
- Aseervatham SB, Suryakala U, Nisha D, Sundaram S, Bose PC, Sivasudha T. Expression pattern of NMDA receptors reveals antiepileptic potential of apigenin 8-C-glucoside and chlorogenic acid in pilocarpine induced epileptic mice. Biomedicine & Pharmacotherapy. 2016; 82: 54-64.
- Szwajgier D, Borowiec K, Pustelniak K. The neuroprotective effects of phenolic acids: Molecular mechanism of action. Nutrients. 2017; 9(5): 477.
- Cevik B, Solmaz V, Aksoy D, Erbas O. Montelukast inhibits pentylenetetrazol-induced seizures in rats. Medical Science Monitor. 2015; 21: 869-74.
- Huang JL, Jing X, Tian X, Qin MC, Xu ZH, Wu DP, et al. Neuroprotective properties of Panax notoginseng saponins via preventing oxidative stress injury in SAMP8 mice. Evidence-Based Complementary and Alternative Medicine. 2017; 2017: 1-8.
- Wilson JX, Gelb AW. Free radicals, antioxidants, and neurologic injury: possible relationship to cerebral protection by anesthetics. Journal of Neurosurgical Anesthesiology. 2002; 14(1): 66-79.
- Ataie A, Ataee R, Shadifar M. Polyphenolic antioxidants and neuronal regeneration. Basic and Clinical Neuroscience. 2016; 7(2): 81-90.
- Agunloye OM, Oboh G. Caffeic acid and chlorogenic acid: Evaluation of antioxidant effect and inhibition of key enzymes linked with hypertension. Journal of Food Biochemistry. 2018; 42(4): e12541.
- Akila P, Vennila L. Chlorogenic acid a dietary polyphenol attenuates isoproterenol induced myocardial oxidative stress in rat myocardium: An in vivo study. Biomedicine & Pharmacotherapy. 2016; 84: 208-14.
- Meng S, Cao J, Feng Q, Peng J, Hu Y. Roles of chlorogenic acid on regulating glucose and lipids metabolism: a review. Evidence-Based Complementary and Alternative Medicine. 2013; 2013.
- Naveed M, Hejazi V, Abbas M, Kamboh AA, Khan GJ, Shumzaid M, et al. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomedicine & Pharmacotherapy. 2018; 97:67-74.
- Ambrogini P, Torquato P, Bartolini D, Albertini MC, Lattanzi D, Di Palma M, et al. Excitotoxicity, neuroinflammation and oxidant stress as molecular bases of epileptogenesis and epilepsy-derived neurodegeneration: The role of vitamin E. Biochimica et Biophysica Acta (BBA)-Molecular Basis of Disease. 2019; 1865(6): 1098-112.
- Paredes RG, Ågmo A. GABA and behavior: the role of receptor subtypes. Neuroscience & Biobehavioral Reviews. 1992; 16(2): 145-70.
- Binder DK. The role of BDNF in epilepsy and other diseases of the mature nervous system. In: Binder DK, Scharfman HE, editors. Recent advances in epilepsy research. Advances in experimental medicine and biology. vol 548. Boston: Springer; 2004. p. 34-56.
- Palma E, Torchia G, Limatola C, Trettel F, Arcella A, Cantore G, et al. BDNF modulates GABAA receptors microtransplanted from the human epileptic brain to Xenopus oocytes. Proceedings of the National Academy of Sciences. 2005; 102(5): 1667-72.
Type of Study: Research |
Subject:
General
Received: 2018/10/7 | Accepted: 2021/10/19 | Published: 2021/12/19
Received: 2018/10/7 | Accepted: 2021/10/19 | Published: 2021/12/19
References
1. Hernández-Ronquillo L, Adams S, Ballendine S, Téllez-Zenteno JF. Epilepsy in an elderly population: Classification, etiology and drug resistance. Epilepsy Research. 2018; 140: 90-4.
2. Erdő F, Denes L, de Lange E. Age-associated physiological and pathological changes at the blood–brain barrier: a review. Journal of Cerebral Blood Flow & Metabolism. 2017; 37(1): 4-24.
3. Mercado NM, Collier TJ, Sortwell CE, Steece-Collier K. BDNF in the aged brain: translational implications for Parkinson’s disease. Austin Neurology & Neurosciences. 2017; 2(2).
4. Mariga A, Mitre M, Chao MV. Consequences of brain-derived neurotrophic factor withdrawal in CNS neurons and implications in disease. Neurobiology of Disease. 2017; 97: 73-9.
5. Brunton L, Chabner B, Knollman B. Goodman and Gilman's the pharmacological basis of therapeutics. New York: McGraw-Hill; 1996.
6. Bethmann K, Fritschy JM, Brandt C, Löscher W. Antiepileptic drug resistant rats differ from drug responsive rats in GABAA receptor subunit expression in a model of temporal lobe epilepsy. Neurobiology of Disease. 2008; 31(2): 169-87.
7. Ali I, Chugh D, Ekdahl CT. Role of fractalkine–CX3CR1 pathway in seizure-induced microglial activation, neurodegeneration, and neuroblast production in the adult rat brain. Neurobiology of Disease. 2015; 74:194-203.
8. Bravo L. Polyphenols: chemistry, dietary sources, metabolism, and nutritional significance. Nutrition Reviews. 1998; 56(11): 317-33.
9. Basli A, Soulet S, Chaher N, Mérillon JM, Chibane M, Monti JP, et al. Wine polyphenols: potential agents in neuroprotection. Oxidative Medicine and Cellular Longevity. 2012; 2012: 1-14.
10. Ozcan T, Akpinar-Bayizit A, Yilmaz-Ersan L, Delikanli B. Phenolics in human health. International Journal of Chemical Engineering and Applications. 2014; 5(5): 393-6.
11. Aseervatham SB, Suryakala U, Nisha D, Sundaram S, Bose PC, Sivasudha T. Expression pattern of NMDA receptors reveals antiepileptic potential of apigenin 8-C-glucoside and chlorogenic acid in pilocarpine induced epileptic mice. Biomedicine & Pharmacotherapy. 2016; 82: 54-64.
12. Szwajgier D, Borowiec K, Pustelniak K. The neuroprotective effects of phenolic acids: Molecular mechanism of action. Nutrients. 2017; 9(5): 477.
13. Cevik B, Solmaz V, Aksoy D, Erbas O. Montelukast inhibits pentylenetetrazol-induced seizures in rats. Medical Science Monitor. 2015; 21: 869-74.
14. Huang JL, Jing X, Tian X, Qin MC, Xu ZH, Wu DP, et al. Neuroprotective properties of Panax notoginseng saponins via preventing oxidative stress injury in SAMP8 mice. Evidence-Based Complementary and Alternative Medicine. 2017; 2017: 1-8.
15. Wilson JX, Gelb AW. Free radicals, antioxidants, and neurologic injury: possible relationship to cerebral protection by anesthetics. Journal of Neurosurgical Anesthesiology. 2002; 14(1): 66-79.
16. Ataie A, Ataee R, Shadifar M. Polyphenolic antioxidants and neuronal regeneration. Basic and Clinical Neuroscience. 2016; 7(2): 81-90.
17. Agunloye OM, Oboh G. Caffeic acid and chlorogenic acid: Evaluation of antioxidant effect and inhibition of key enzymes linked with hypertension. Journal of Food Biochemistry. 2018; 42(4): e12541.
18. Akila P, Vennila L. Chlorogenic acid a dietary polyphenol attenuates isoproterenol induced myocardial oxidative stress in rat myocardium: An in vivo study. Biomedicine & Pharmacotherapy. 2016; 84: 208-14.
19. Meng S, Cao J, Feng Q, Peng J, Hu Y. Roles of chlorogenic acid on regulating glucose and lipids metabolism: a review. Evidence-Based Complementary and Alternative Medicine. 2013; 2013.
20. Naveed M, Hejazi V, Abbas M, Kamboh AA, Khan GJ, Shumzaid M, et al. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomedicine & Pharmacotherapy. 2018; 97:67-74.
21. Ambrogini P, Torquato P, Bartolini D, Albertini MC, Lattanzi D, Di Palma M, et al. Excitotoxicity, neuroinflammation and oxidant stress as molecular bases of epileptogenesis and epilepsy-derived neurodegeneration: The role of vitamin E. Biochimica et Biophysica Acta (BBA)-Molecular Basis of Disease. 2019; 1865(6): 1098-112.
22. Paredes RG, Ågmo A. GABA and behavior: the role of receptor subtypes. Neuroscience & Biobehavioral Reviews. 1992; 16(2): 145-70.
23. 23. Binder DK. The role of BDNF in epilepsy and other diseases of the mature nervous system. In: Binder DK, Scharfman HE, editors. Recent advances in epilepsy research. Advances in experimental medicine and biology. vol 548. Boston: Springer; 2004. p. 34-56.
24. Palma E, Torchia G, Limatola C, Trettel F, Arcella A, Cantore G, et al. BDNF modulates GABAA receptors microtransplanted from the human epileptic brain to Xenopus oocytes. Proceedings of the National Academy of Sciences. 2005; 102(5): 1667-72.
Send email to the article author
| Rights and permissions | |
 |
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License. |
